Embora todas as características da planta de ervilha de Mendel tenham se comportado de acordo com a lei do sortimento independente, sabemos agora que algumas combinações de alelos não são herdadas independentemente umas das outras. Os genes que estão localizados em cromossomos separados e não homólogos sempre serão classificados independentemente.
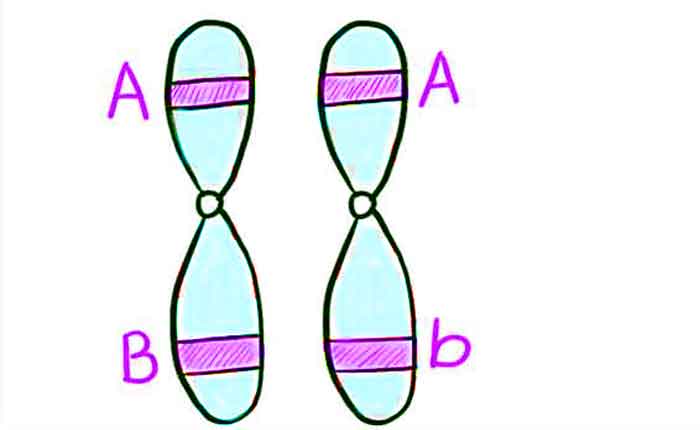
Os cromossomos homólogos possuem os mesmos genes na mesma ordem, embora os alelos específicos do gene possam ser diferentes em cada um dos dois cromossomos. Lembre-se de que, durante a interfase e a prófase I da meiose, os cromossomos homólogos primeiro se replicam e depois fazem sinapses, com genes semelhantes nos homólogos se alinhando entre si.
Nesta fase, segmentos de cromossomos homólogos trocam segmentos lineares de material genético ( Figura ). Este processo é chamado de recombinação, ou crossover, e é um processo genético comum. Como os genes estão alinhados durante a recombinação, a ordem dos genes não é alterada. Em vez disso, o resultado da recombinação é que os alelos maternos e paternos são combinados no mesmo cromossomo. Através de um determinado cromossomo, vários eventos de recombinação podem ocorrer, causando extenso deslocamento dos alelos.

Quando dois genes estão localizados no mesmo cromossomo, eles são considerados ligados, e seus alelos tendem a ser transmitidos através da meiose juntos. Para exemplificar isso, imagine um cruzamento diíbrido envolvendo a cor da flor e a altura da planta em que os genes estão próximos um do outro no cromossomo.
Se um cromossomo homólogo tem alelos para plantas altas e flores vermelhas, e o outro cromossomo possui genes para plantas curtas e flores amarelas, então quando os gametas são formados, os alelos altos e vermelhos tenderão a se juntar em um gameta e o curto e alelos amarelos irão para outros gametas. Estes são chamados de genótipos parentais porque foram herdados intactos dos pais dos gametas produtores individuais.
Mas ao contrário, se os genes estivessem em cromossomos diferentes, não haverá gametas com alelos altos e amarelos e sem gametas com alelos curtos e vermelhos. Se você criar um quadrado de Punnett com esses gametas, verá que a previsão mendeliana clássica de um resultado 9: 3: 3: 1 de uma cruz diíbrida não se aplica.
A medida que a distância entre dois genes aumenta, a probabilidade de um ou mais cruzamentos entre eles aumenta e os genes se comportam mais como se estivessem em cromossomos separados. Os geneticistas usaram a proporção de gametas recombinantes (aqueles que não são como os pais) como uma medida de quão distantes estão os genes de um cromossomo.
Usando essas informações, eles construíram mapas de ligação de genes em cromossomos para organismos bem estudados, incluindo seres humanos. 1 resultado de um cruzamento diíbrido não se aplicaria. À medida que a distância entre dois genes aumenta, a probabilidade de um ou mais cruzamentos entre eles aumenta e os genes se comportam mais como se estivessem em cromossomos separados.
Os geneticistas usaram a proporção de gametas recombinantes (aqueles que não são como os pais) como uma medida de quão distantes estão os genes de um cromossomo. Usando essas informações, eles construíram mapas de ligação de genes em cromossomos para organismos bem estudados, incluindo seres humanos. 1 resultado de um cruzamento diíbrido não se aplicaria.
A medida que a distância entre dois genes aumenta, a probabilidade de um ou mais cruzamentos entre eles aumenta e os genes se comportam mais como se estivessem em cromossomos separados. Os geneticistas usaram a proporção de gametas recombinantes (aqueles que não são como os pais) como uma medida de quão distantes estão os genes de um cromossomo. Usando essas informações, eles construíram mapas de ligação de genes em cromossomos para organismos bem estudados, incluindo seres humanos.
A publicação seminal de Mendel não faz menção à ligação, e muitos pesquisadores questionaram se ele encontrou ligação, mas optou por não publicar esses cruzamentos por preocupação de que eles invalidariam seu postulado de sortimento independente.
A ervilha tem sete cromossomos e alguns sugeriram que a escolha de sete características não foi uma coincidência. No entanto, mesmo que os genes que ele examinou não estivessem localizados em cromossomos separados, é possível que ele simplesmente não tenha observado a ligação por causa dos extensos efeitos da recombinação.
Veja também:
Epistasia
Os estudos de Mendel em plantas de ervilha implicavam que a soma do fenótipo de um indivíduo era controlada por genes (ou como ele os chamava, fatores unitários), de modo que cada característica era distinta e completamente controlada por um único gene.
De fato, características observáveis únicas estão quase sempre sob a influência de múltiplos genes (cada um com dois ou mais alelos) atuando em uníssono. Por exemplo, pelo menos oito genes contribuem para a cor dos olhos em humanos.
Em alguns casos, vários genes podem contribuir para aspectos de um fenótipo comum sem que seus produtos genéticos interajam diretamente. No caso do desenvolvimento de órgãos, por exemplo, os genes podem ser expressos sequencialmente, com cada gene aumentando a complexidade e especificidade do órgão.
Os genes podem funcionar de forma complementar ou sinérgica, de modo que dois ou mais genes expressos simultaneamente afetam um fenótipo. Um exemplo aparente disso ocorre com a cor da pele humana, que parece envolver a ação de pelo menos três (e provavelmente mais) genes. Casos em que a herança de uma característica como cor da pele ou altura humana dependem dos efeitos combinados de numerosos genes são chamados de herança poligênica.
Os genes também podem se opor, com um gene suprimindo a expressão de outro. Na epistasia , a interação entre os genes é antagônica, de modo que um gene mascara ou interfere na expressão do outro. “Epistasia” é uma palavra composta de raízes gregas que significa “ficar em pé”. Diz-se que os alelos que estão sendo mascarados ou silenciados são hipostáticos para os alelos epistáticos que estão fazendo o mascaramento. Muitas vezes, a base bioquímica da epistasia é uma via genética na qual a expressão de um gene é dependente da função de um gene que o precede ou segue na via.
Um exemplo de epistasia é a pigmentação em camundongos. A cor da pelagem do tipo selvagem, cutia (AA) é dominante para pêlo de cor sólida (aa). No entanto, um gene C separado, quando presente como o homozigoto recessivo (cc), nega qualquer expressão de pigmento do gene A e resulta em um camundongo albino ( Figura ). Portanto, os genótipos AAcc , Aacc e aaccproduzem o mesmo fenótipo albino. Um cruzamento entre heterozigotos para ambos os genes ( AaCc x AaCc ) geraria descendentes com uma razão fenotípica de 9 agouti: 3 pretos: 4 albinos ( Figura ). Neste caso, o gene C é epistático ao gene A.

Os alelos nem sempre se comportam em padrões dominantes e recessivos. A dominância incompleta descreve situações em que o heterozigoto exibe um fenótipo intermediário entre os fenótipos homozigotos. A codominância descreve a expressão simultânea de ambos os alelos no heterozigoto. Embora os organismos diploides só possam ter dois alelos para um dado gene, é comum que mais de dois alelos para um gene existam em uma população.
Em humanos, como em muitos animais e algumas plantas, as fêmeas têm dois cromossomos X e os machos têm um cromossomo X e um Y. Os genes que estão presentes no cromossomo X, mas não no cromossomo Y, são ligados ao cromossomo X, de modo que os machos só herdam um alelo para o gene e as fêmeas herdam dois.
De acordo com a lei de Mendel do sortimento independente, os genes se classificam independentemente uns dos outros em gametas durante a meiose. Isso ocorre porque os cromossomos, nos quais os genes residem, variam independentemente durante a meiose e os cruzamentos fazem com que a maioria dos genes nos mesmos cromossomos também se comporte independentemente.
Quando os genes estão localizados próximos ao mesmo cromossomo, seus alelos tendem a ser herdados juntos. Isso resulta em índices descendentes que violam a lei de Mendel de sortimento independente. No entanto, a recombinação serve para trocar material genético em cromossomos homólogos, de modo que os alelos maternos e paternos possam ser recombinados no mesmo cromossomo.
É por isso que os alelos de um determinado cromossomo nem sempre são herdados juntos. A recombinação é um evento aleatório que ocorre em qualquer parte de um cromossomo. Assim sendo,
Independentemente de estarem ou não ordenando independentemente, os genes podem interagir no nível dos produtos gênicos, de modo que a expressão de um alelo para um gene mascara ou modifica a expressão de um alelo para um gene diferente. Isso é chamado de epistasia.
Notas de rodapé
- 1 Sumiti Vinayak et al., “Origem e Evolução do Plasmodium falciparum Resistente à Sulfadoxina ”, PLoS Pathogens 6 (2010): e1000830.
- https://openoregon.pressbooks.pub/mhccbiology102/chapter/12-9-linked-genes-violate-the-law-of-independent-assortment/
Glossário
- codominância
- em um heterozigoto, expressão completa e simultânea de ambos os alelos para a mesma característica
- epistasia
- uma interação entre genes de tal forma que um gene mascara ou interfere com a expressão de outro
- hemizigoto
- a presença de apenas um alelo para uma característica, como na ligação X; hemizigosidade faz descrições de dominância e recessividade irrelevantes
- dominância incompleta
- em um heterozigoto, expressão de dois alelos contrastantes, de modo que o indivíduo exiba um fenótipo intermediário
- ligação
- um fenômeno no qual os alelos que estão localizados próximos uns dos outros no mesmo cromossomo são mais propensos a serem herdados juntos
- recombinação
- o processo durante a meiose em que os cromossomos homólogos trocam segmentos lineares de material genético, aumentando assim drasticamente a variação genética na prole e separando genes ligados
- tipo selvagem
- genótipo ou fenótipo de ocorrência mais comum para uma dada característica encontrada em uma população
- Vinculado ao X
- um gene presente no cromossomo X, mas não no cromossomo Y